Research Weisser Group
We are interested in the ecology of species interactions and in the consequences of these interactions at the population, community and ecosystem level. Most of the work focuses on the multi-trophic interactions between plants, herbivorous insects and their natural enemies, but some studies involve plant-decomposer, plant-pollinator and plant-seed disperser interactions. Aphids, grasshoppers, bees, but also beetles, springtails, earthworms and slugs are common model organisms.
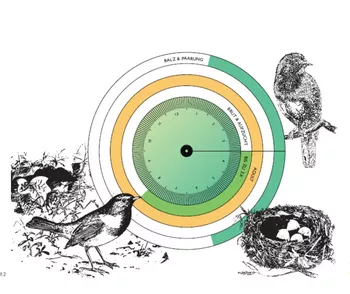
Animal-Aided Design – bridging the gap between landscape architecture and conservation
Biodiversity underlies many of the ecosystem services demanded by humans. To maintain biodiversity and ecosystem services, the creation of a ‘green infrastructure’ has been proposed for Europe. The basic idea is that this green infrastructure is as important for the human society as other infrastructures such as the electricity grid.
It is unclear, however, how such a green infrastructure should be created. Urban open spaces are currently planned by landscape architects with a primary focus on aesthetic design, or by city planners with little expertise in ecology. As a consequence, standard planning procedures do not create a green infrastructure. On the other hand, conservation often targets the few remaining areas with little influence of humans, also in cities. While this will conserve biodiversity and maintain certain ecosystem services, it also does not represent a targeted planning process aimed at providing a green infrastructure also in places where there are no wilderness areas left. In fact, conservation and urban planning often work against one another rather than together, for example when a planning process is executed with little reference to biodiversity and when nature protection laws interfere with this planning process by requiring adaptation of the design due to the occurrence of a protected species.
We have developed Animal-Aided Design® as a methodology for the design of open spaces that can help to overcome this difference between landscape architecture and conservation. The basic idea of Animal-Aided Design® (in short, AAD) is to include the presence of animals in the planning process, such that they are an integral part of the design. For AAD, the desired species are chosen at the beginning of a project. The requirements of the target species, i.e. their life-cycles, then set boundary conditions and serve as an inspiration for the design.
Download booklet that describes Animal-Aided Design (only in German). Please use the two-pages view in Adobe Acrobat!
Animal-Aided Design is a cooperation project with Dr. Thomas Hauck, Universität Kassel Link to Polinna Hauck Landscape+Urbanism
and a subproject of the Centre for Urban Ecology and Climate Adaptation (ZSK).
Mechanisms underlying the relationship between biodiversity and ecosystem functioning

In the past two decades, an increasing number of studies has been performed to analyze the relationship between biodiversity and ecosystem functioning. We contribute to the search for ecosystem consequences of biodiversity loss in the DFG-funded Jena Experiment, a grassland biodiversity experiment coordinated by our research group, and in the BMBF-funded BIOLOG DIVA-Jena project (completed at the end of 2010), a study in semi-natural grasslands along a plant species richness gradient. A major conclusion that can be drawn from this research is that a low diversity in an assemblage is often associated with a lowered mean (and an increase in the variance) in many of the ecosystem variables investigated. In the Jena Experiment, for example, about 40% of ecosystem variables are affected by plant species richness. One major shortcoming of the current state of knowledge is, however, that for many of the observed biodiversity effects we do not know the underlying mechanism. We are interested in how species interaction change with increasing plant species richness and how these changes underlie the observed biodiversity effects on ecosystem variables. See the descriptions of the Jena Experiment and BIOLOG Diva-Jena for more detail.
Land-use, biodiversity and ecosystem functioning
A major driver of biodiversity loss is land use change. Human land use not only changes biodiversity, but also ecosystem variables either directly or via the change in biodiversity. In a number of projects we investigate the feedback loops between land use, biodiversity change and ecosystem variables, focusing on the role of species interactions. In the DFG-funded Biodiversity Exploratories project, the focus is on land use in forest and grassland ecosystems in Germany. This project represents a major cooperation effort of more than 40 research groups.
In the completed project Degradation of South Brazilian grasslands, we cooperated with the restoration ecology group at TUM and colleagues at the Universidade Estadual do Rio Grande do Sul in Porto Alegre, Brasil, to investigate biodiversity and ecosystem functions in differently degraded grasslands. The natural grasslands in south Brazil have at a large scale been converted into tree plantations or agricultural land or are affected by by burning, grazing and seeding with alien species. See the following publications for the outcome:
| Leidinger, J., M. M. Gossner, W. W. Weisser, C. Koch, Z. L. R. Cayllahua, L. R. Podgaiski, M. M. Duarte, A. S. F. d. Araújo, J. Hermann, J. Kollman and S. T. Meyer. 2017. Historic and recent land use affects ecosystem functions in subtropical grasslands in southern Brazil. Ecosphere 8:e02032. PDF |
| Koch, C., T. Conradi, M. M. Gossner, J. M. Hermann, J. Leidinger, S. T. Meyer, G. E. Overbeck, W. W. Weisser and J. Kollmann. 2016. Management intensity and temporary conversion to other land-use types affect plant diversity and species composition of subtropical grasslands in southern Brazil. Applied Vegetation Science 19:589-599. |
Former projects on this topic include Agripopes, funded by DFG and the European Science Foundation, which investigated AGRIcultural POlicy-induced changes in agricultural landscapes and their effects on biodiversity and ecosystem services.
In project recently funded by the Bavarian state forest administration, we evaluate a dead-wood enrichment strategy that aims to integrate conservation with economic forestry. The strategy was implemented in beech forests in the Steigerwald forest district in Bavaria almost 10 years ago and we investigate whether a) there has been a measurable increase in deadwood in the forest stands, and b) whether an enrichment in deadwood has resulted in a measurable increase in deadwood-associated biodiversity.
Plant-insect metacommunity ecology and genetics

In an ecological community, species interactions structure local populations and influence species coexistence. Two different, yet complementary, approaches to studying species interactions are community genetics, which recognises that within-species genetic variation can alter the interactions between different species; and, metacommunity ecology, which emphasizes the role dispersal plays in structuring local communities.
We investigate how community genetic interactions influence metacommunity dynamics, using a model tansy (Tanacetum vulgare) plant – aphid system. Individual tansy plants act as 'islands of resource' and, on this plant, classical metapopulation structuring is exhibited by competing specialist aphids and their parasitoids (e.g. We study this system using field, common garden and greenhouse experiments to uncover the mechanisms driving metapopulation and metacommunity dynamics. We also examine aphid population genetic structure and the effect of plant genetic variation.
One focus has been for several years the role of predation for prey population ecology and genetics. In 1999, we demonstrated that pea aphids, Acyrthosiphon pisum, produce winged offspring when exposed to predatory ladybirds, the first example of an enemy-induced morphological change facilitating dispersal (Weisser et al. 1999). We continue investigating the role of natural enemies for aphid wing induction, and have found wing-inducing effects of e.g. aphid parasitoids (Sloggett & Weisser 2002), lacewing larvae (Kunert & Weisser 2003) and aphid entomopathogenic fungi (Hatano et al. 2012). We also investigate the underlying mechanism of this response to the presence of natural enemies. Particular attention is paid to the role of (E)-ß-Farnesene (EBF), the main component of aphid alarm pheromone. We have shown (Kunert et al. 2005) that wing induction in the presence of natural enemies is mediated by EBF. Currently, we focus on the role that EBF plays in the network of ecological interactions between plants, aphids, ants and the natural enemies of aphids using e.g. rapid gas chromatography for real-time analysis of volatile communication (Joachim et al. 2013, 2014).
In our current DFG-funded project on tansy aphids metacommunity genetics we cooperate closely with the Helmholtz Zentrum München, who study the plant chemical ecology, and investigate the effect of expression of mono- and sesquiterpenes on metacommunity dynamics.
Joachim, C., I. Vosteen and W. W. Weisser. 2014. The aphid alarm pheromone (E)-beta-farnesene does not act as a cue for predators searching on a plant. Chemoecology in press.
Joachim, C. and W. W. Weisser. 2013. Real-Time Monitoring of (E)-beta-Farnesene Emission in Colonies of the Pea Aphid, Acyrthosiphon pisum, Under Lacewing and Ladybird Predation. Journal of Chemical Ecology 39:1254-1262. PDF
Hatano, E., J. Baverstock, G. Kunert, J. K. Pell and W. W. Weisser. 2012. Entomopathogenic fungi stimulate transgenerational wing induction in pea aphids, Acyrthosiphon pisum (Hemiptera: Aphididae). Ecological Entomology 37:75-82. PDF
Kunert, G., S. Otto, U. S. R. Rose, J. Gershenzon and W. W. Weisser. 2005. Alarm pheromone mediates production of winged dispersal morphs in aphids. Ecology Letters 8:596-603. PDF
Kunert, G. and W. W. Weisser. 2003. The interplay between density- and trait-mediated effects in predator-prey interactions: a case study in aphid wing polymorphism. Oecologia 135:304-312. PDF
Sloggett, J. J. and W. W. Weisser. 2002. Parasitoids induce production of the dispersal morph of the pea aphid, Acyrthosiphon pisum. Oikos 98:323-333. PDF
Weisser, W. W., C. Braendle and N. Minoretti. 1999. Predator-induced morphological shift in the pea aphid. Proceedings of the Royal Society of London Series B-Biological Sciences 266:1175-1181. PDF
Schwammspinner - Projekt

Technische Universität München, LWF, Universität Würzburg
Studie zu forstlichen und ökologischen Folgen bei Massenvermehrungen des Schwammspinners in Bayern
Die Bekämpfung des Schwammspinners in Eichenmischwäldern ist in jedem Jahr einer Massenvermehrung Gegenstand einer öffentlichen Diskussion. Einerseits können hohe Dichten der Schwammspinnerraupen zu Kahlfraß an Eiche und auch an anderen Baumarten führen. Dieser soll durch eine rechtzeitige Bekämpfung vermieden werden, um Waldbesitzer vor Schäden bis hin zu Bestandeszerstörungen zu bewahren. Andererseits sind Eichenmischwälder sehr artenreich und es kommen in ihnen eine Vielzahl bedrohter Tag- und Nachtfalter, Vögel und andere Arten vor, so dass die Sorge besteht, dass diese kurz- oder auch längerfristig unter einem Insektizideinsatz leiden. Allerdings hat auch der Schwammspinnerausbruch selbst negative Konsequenzen für andere Arten in einem Eichenwald, da die Artengemeinschaft für einige Zeit von nur einer Art, dem Schaderreger, dominiert wird. Während die Biologie des Schwammspinners bekannt ist und es bereits viele gute Studien zur Wirkung einer Entlaubung auf die Eiche gibt, gibt es doch einige Wissensdefizite, die für ein Risikomanagement wichtig sind, z.B. in Bezug auf kurz- und längerfristige Effekte des Schwammspinners auf das Eichenwachstum in Abhängigkeit des Standorts. Auch gibt es bisher noch zu wenige Studien, die die kurz- und längerfristigen Auswirkungen auf Nichtzielorganismen zwischen unbekämpften Flächen (mit hoher Schwammspinnerdichte) und bekämpften Flächen (mit reduzierter Schwammspinnerdichte) vergleichen. Schließlich sind einige internationale Ergebnisse nicht einfach auf Bayern zu übertragen. Die Wechselwirkungen zwischen Schwammspinnerdichten, standörtlichen Gegebenheiten, dem gewählten Management und den Reaktionen der Eichenwälder und ihrer Lebensgemeinschaften können dabei nur in Jahren einer Massenvermehrung untersucht werden, wie sie für die nächsten Jahre erwartet wird.